
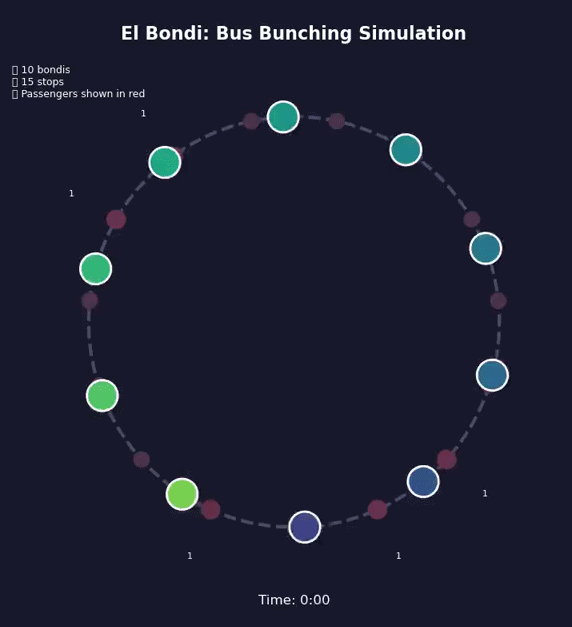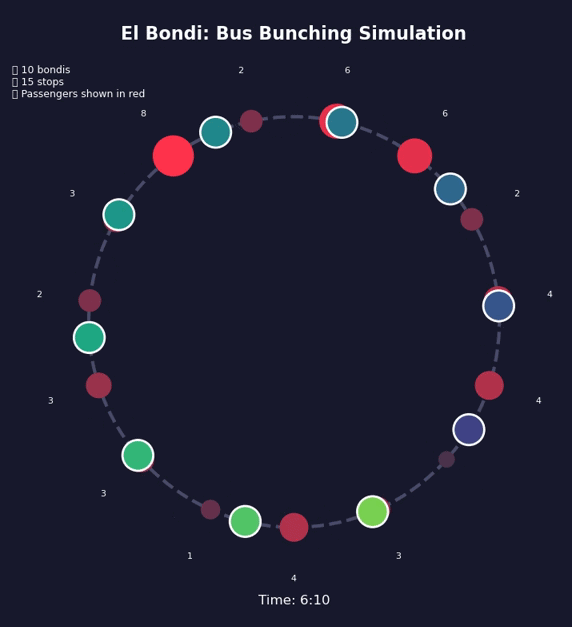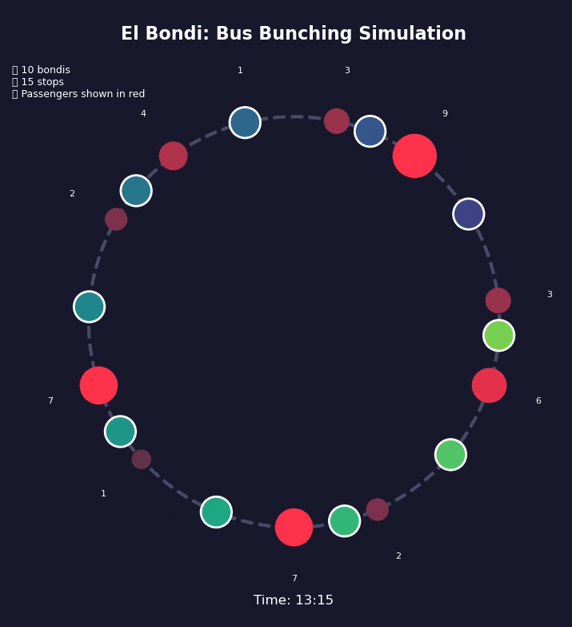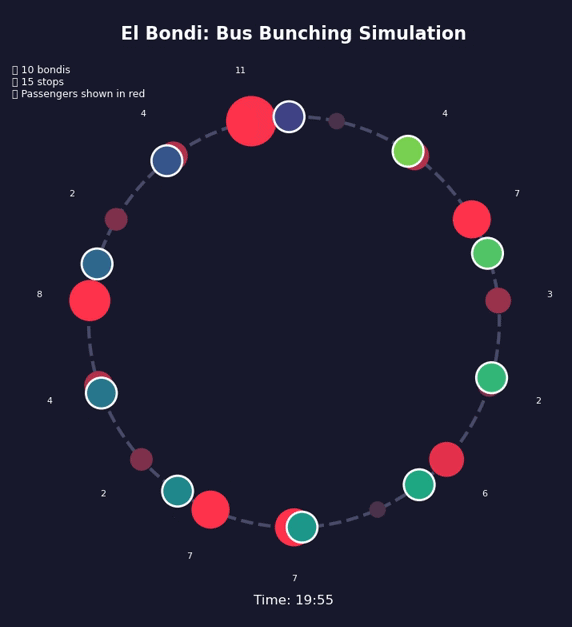
Bondi bunching

During my doctoral years at the University of Buenos Aires, I took the 160 bus every day to get to the office. I spent countless hours at bus stops, witnessing a maddening yet intriguing pattern: I’d wait 20, sometimes 30 minutes without a single bus, and then—suddenly—three would arrive together.
At first, I blamed the drivers, the bus company, or the traffic. What were they doing—waiting to travel together and chat at red lights? Was I just unlucky? It didn’t add up. The more I thought about it, the more I realized it wasn’t random at all, but inevitable. Bus bunching isn’t a failure of planning; it’s the outcome of physics and feedback loops.
A positive feedback loop#
Imagine Bus A hits traffic or finds more passengers than usual at a stop. It falls slightly behind schedule. Now the next stop has had more time to accumulate passengers, so Bus A takes even longer to board them. It falls further behind. Meanwhile, Bus B behind it finds fewer passengers at each stop—it speeds up and catches Bus A.
Traffic, weather, accidents—any event that delays a bus will push it toward this outcome. The gaps between buses don’t correct themselves; they grow.
This is a positive feedback loop. Evenly-spaced buses are an unstable equilibrium. Bus bunching is the stable one. Left alone, bunching is where all buses end up.
So why don’t we see it all the time?
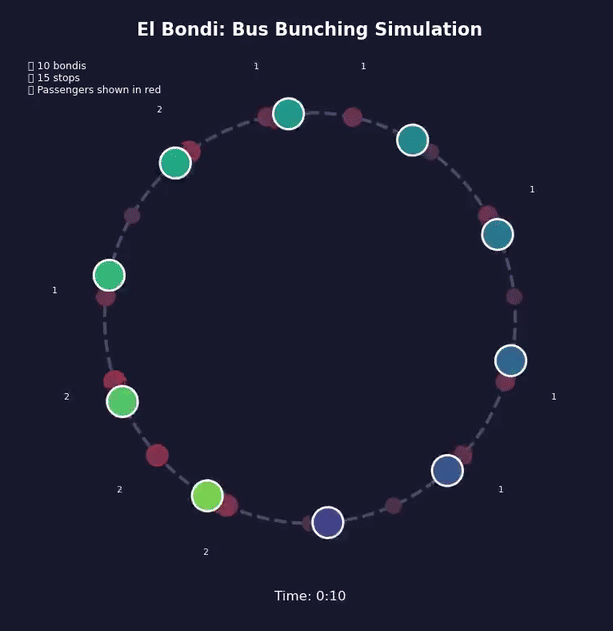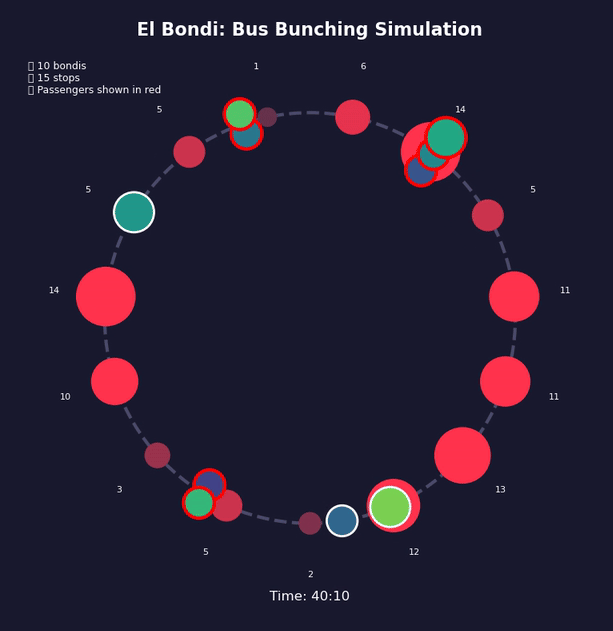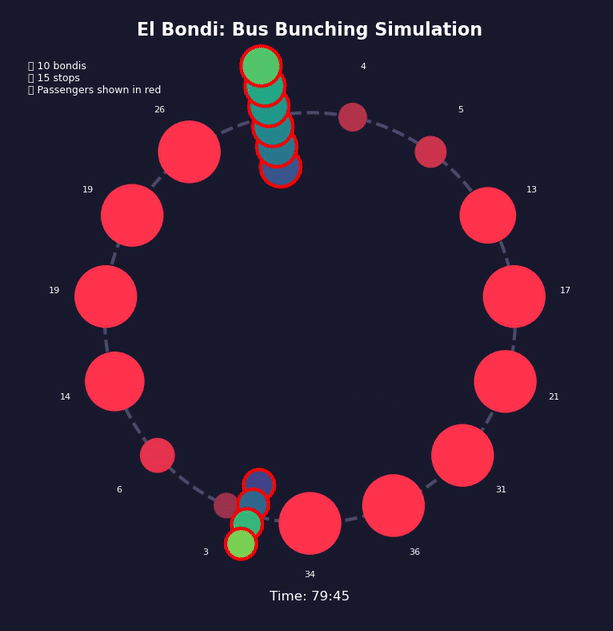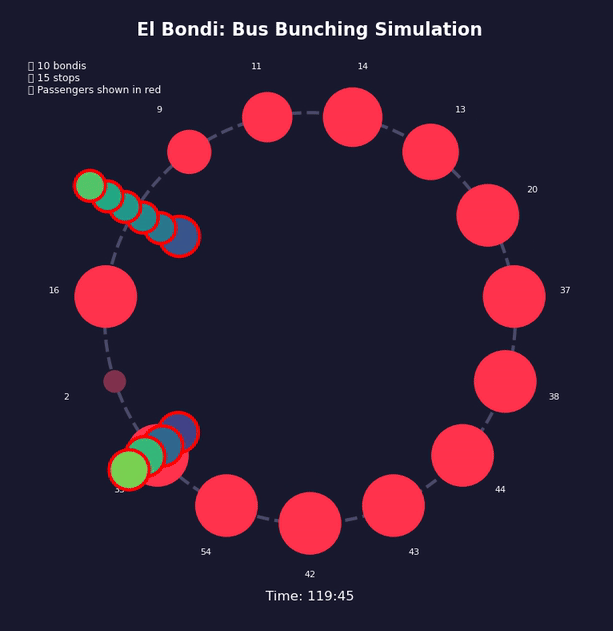
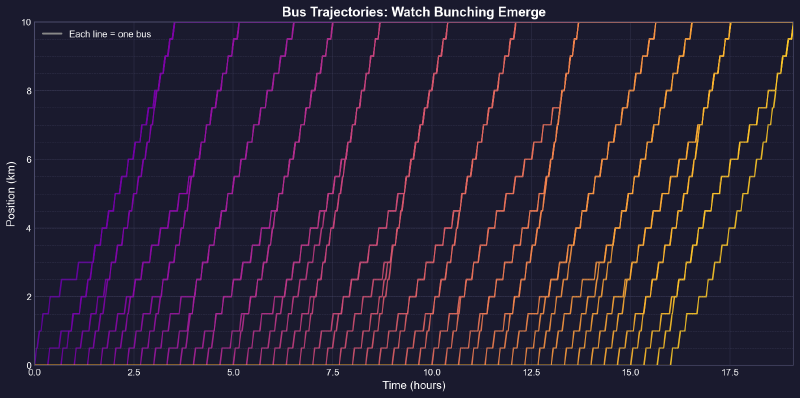
Bus bunching makes service unpredictable: passengers face long waits followed by multiple buses at once, some packed and others nearly empty. It also wastes resources—several large buses traveling together clog traffic while leaving gaps in service elsewhere. The problem gets worse with longer routes, higher frequency, more passengers, unpredictable traffic, and slow boarding—all of which give the feedback loop more room to grow. Transit agencies fight back by holding early buses at stops, padding schedules with extra recovery time, using GPS to adjust speeds in real time, and reducing boarding variability with prepaid fares and multiple doors. Dedicated bus lanes help by removing traffic randomness. None of these eliminate bunching, but they slow it down.
Bunching Beyond Buses#
The same instability that bunches buses appears throughout nature. The ingredients are simple: units moving in sequence along a shared path, where the speed of each unit depends on local conditions. Whenever these conditions hold, bunching emerges.
Ribosomes translating mRNA bunch up behind slow codons. Molecular motors walking along microtubules cluster when one stalls. Ants following pheromone trails pile up on popular routes. Cars on a highway form the stop-and-go waves we call traffic jams. The mathematics underlying all these systems is remarkably similar—a positive feedback loop that amplifies small perturbations into large clusters.
Sometimes bunching is a problem to solve. Sometimes it’s a feature to exploit.
Cell Bunching in Cancer Metastasis#
In cancer metastasis, tumor cells escape and invade surrounding tissue. Clinically, it is well established that cells travel from tumors as clusters, and these clusters are far more dangerous than single cells—they survive better in the bloodstream and are more likely to seed new tumors.
But why do cells cluster in the first place?
As a cell moves through the extracellular matrix toward blood vessels, it degrades the matrix and leaves behind a softened, remodeled track—a footprint. Follower cells encounter this footprint and move faster along it. They catch up to the leader. The same feedback loop that bunches buses bunches cells.
The footprint degrades over time, just as passengers accumulate at bus stops over time. Both create a memory effect: the longer since the last bus (or cell) passed, the more “resistance” the next one encounters. In buses, this means more passengers to board. In cells, it means a more intact matrix to degrade.
This memory effect controls whether bunching occurs. If buses run frequently, small delays compound before the system resets—bunching is inevitable. If buses run infrequently, fluctuations in boarding time aren’t enough for one bus to catch another. The same logic applies to cells: high escape rates from the tumor lead to clustering; low escape rates keep cells isolated.
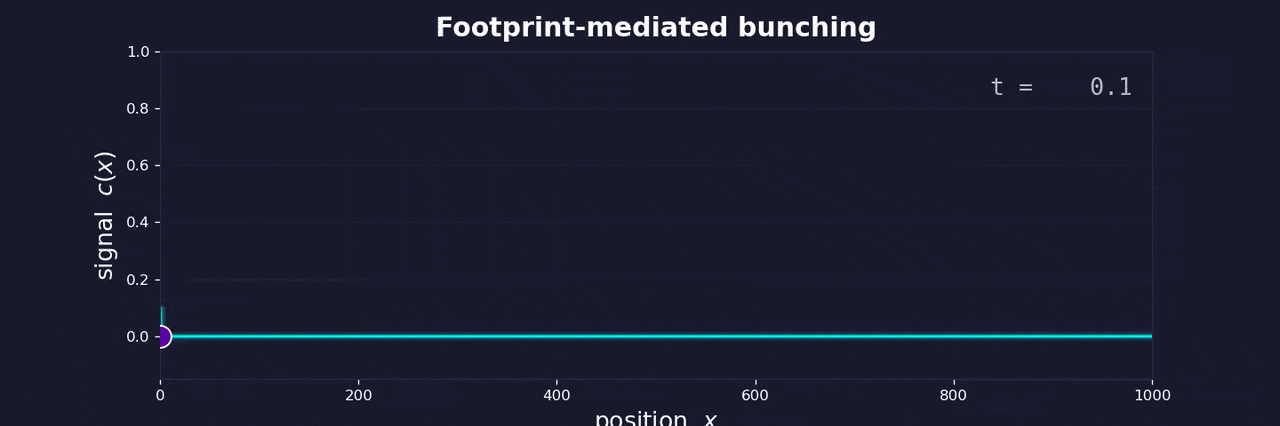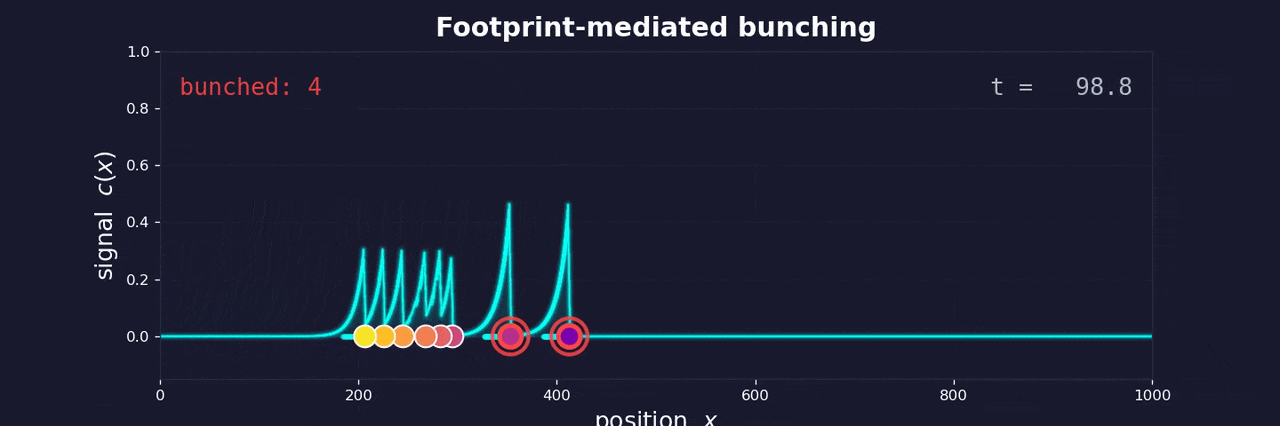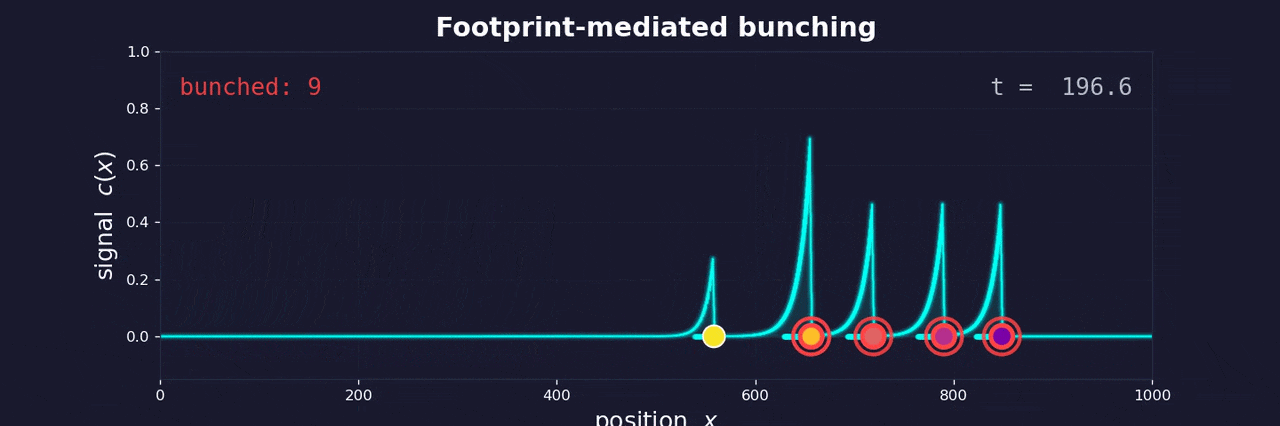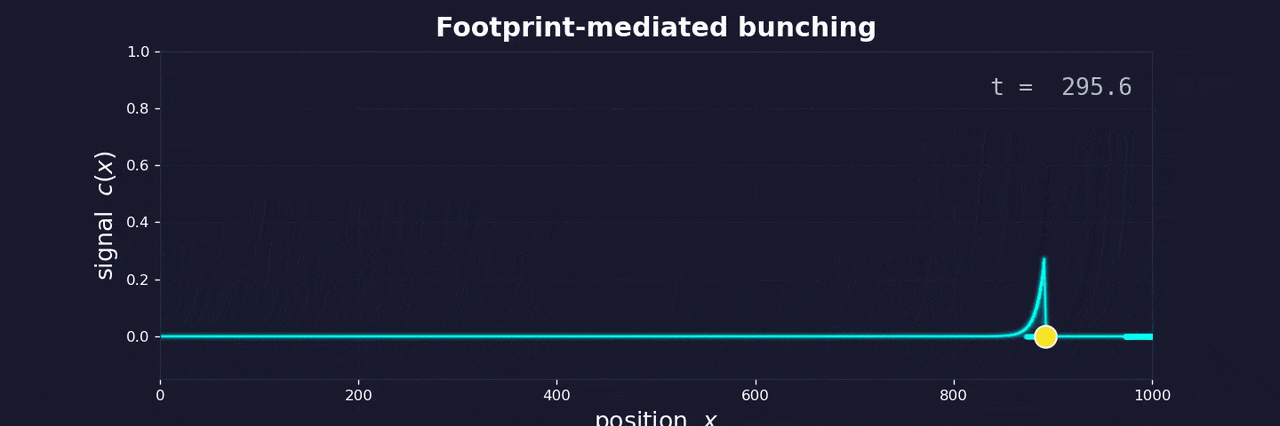
This reframes metastatic clustering not as active coordination between cells, but as an emergent physical phenomenon—the inevitable consequence of cells moving along a shared path with memory. The same instability that frustrated me at Buenos Aires bus stops may help explain why cancer spreads the way it does.
Understanding the physics of cell bunching opens new questions: Can we disrupt the footprint to prevent clustering? Can we tune matrix properties to keep cells isolated? The answers may lie in the same mathematics that transit engineers use to keep buses evenly spaced.
There are no articles to list here yet.